To enhance the functionality of black tea beverages, this study investigated fermentation by eight Lactobacillus plantarum strains in black tea-sugar broth over 48 hours. Key analyses included pH, acidity, organic acids, sugars, phenols, and flavonoids. Strain-specific profiles emerged: some strains showed strong growth and acid production, while others maintained stable pH. Principal component analysis (77.32% variance) highlighted acidification and sugar metabolism as main drivers. Bioactive compounds like phenols and flavonoids remained stable, preserving antioxidant benefits. These insights offer a framework for optimizing fermentation, enabling producers to create probiotic-rich, health-promoting drinks with improved gut health and flavor potential.
The global demand for health-promoting foods has surged in recent years, driven by increasing consumer awareness of diet-related health benefits. Among these, fermented tea beverages, such as kombucha, tea wine, and tea vinegar, have gained prominence due to their unique flavors and rich bioactive compounds, including antioxidants and probiotics 1, 2. These beverages are produced through microbial fermentation, which modifies tea’s natural constituents, enhancing health attributes like oxidative stress reduction and gut health support 3.
Traditionally, fermentation of tea relies on yeast and acetic acid bacteria (AAB), which drive acid accumulation and flavor development. However, recent omics-based studies have highlighted the emerging role of lactic acid bacteria (LAB) in polysaccharide metabolism, acid regulation, and probiotic effects, expanding the microbial diversity in tea fermentation 4, 5. Among LAB, Lactobacillus plantarum stands out for its metabolic versatility and acid tolerance, making it a promising candidate for fermented food applications 6, 7. This strain efficiently utilizes sugars (e.g., sucrose, glucose, fructose) via glycolytic pathways, producing lactic acid and exopolysaccharides (EPS) that improve texture and health value 8, 9. For instance, Zhang et al 10. demonstrated that L. plantarum C88 converts sugars into lactic acid and EPS (LPC-1), reducing sugar content while enhancing gut barrier function through antioxidant activity.
Despite these advances, the specific contributions of L. plantarum to tea fermentation, particularly black tea, remain underexplored. Black tea, rich in polyphenols, poses unique challenges due to its complex matrix and potential inhibition of LAB by dominant AAB 11. Existing studies on kombucha suggest LAB enhances acidity and EPS production 12, but strain-specific effects on sugar metabolism, organic acid profiles, and functional component stability (e.g., polyphenols, flavonoids) in black tea are poorly understood. This knowledge gap hinders the optimization of fermentation processes for functional black tea beverages.
To address this, the present study screened eight L. plantarum strains isolated from food sources to investigate their metabolic behaviors during black tea-sugar broth fermentation. The research focused on physicochemical changes (pH, total acidity, organic acids, sugars) and the dynamics of functional components (total phenols, flavonoids). These findings aim to elucidate L. plantarum’s role in sugar utilization and acidogenesis, while assessing its potential to preserve bioactive compounds, providing a scientific basis for developing high-quality, health-promoting black tea-based beverages.
Black tea (Camellia sinensis var. assamica) was sourced from Yiming Ecological Tea Co., Ltd. (Nanping, China). Sucrose was obtained from Angel Yeast Co., Ltd. (Chongzuo, China). All other reagents, including sodium hydroxide (NaOH), 3,5-dinitrosalicylic acid, phenol, sodium sulfite, potassium sodium tartrate, aluminum chloride (AlCl3), sodium nitrite (NaNO2), sodium carbonate (Na2CO3), Folin-Ciocalteu reagent, and standards for organic acids (formic, citric, malic, lactic, acetic) and carbohydrates (sucrose, fructose, glucose), were purchased from Sinopharm Chemical Reagent Co., Ltd. (Beijing, China). All reagents were of analytical grade unless otherwise specified.
2.2. Fermentation of Black Tea Broth with Lactobacillus plantarumEight L. plantarum strains (HD2, JF2, J10, RU1, SC1, J30, GA6 and LB3), isolated and preserved at the Food Safety and Quality Control Laboratory of Yangzhou University, were used to ferment a sucrose-supplemented black tea broth. The broth was prepared by infusing 5.0 g of black tea leaves in 1 L boiling ultrapure water for 15 min, filtering, cooling to 50°C, adding 50.0 g sucrose (5%, w/v), and sterilizing at 121°C for 15 min. Each strain was revived from glycerol stocks in de Man, Rogosa, and Sharpe (MRS) broth at 37°C for 24 h, subcultured three times, harvested by centrifugation (5000 × g, 10 min, 4°C), and resuspended in saline to approximately 1 × 108 CFU/mL. An aliquot corresponding to 1% (v/v) of this suspension was inoculated into the sterilized tea broth, which was incubated statically at 37°C for 48 h, and samples were withdrawn at 12, 24, 36 and 48 h for subsequent analyses.
2.3. Biochemical AnalysesCell growth was monitored by measuring optical density at 600 nm (OD600) using a WGZ-XT turbidity meter (Hangzhou Qiwei Instrument Co., Ltd., Hangzhou, China). Samples were diluted to an OD600 range of 0.05–1.0, and readings were converted to colony-forming units (CFU/mL) using a pre-established standard curve. The pH was measured with a PHS-3C pH meter (REX Instruments, Shanghai, China). Total titratable acidity (TA) was determined by acid-base titration following the Chinese standard GB 12456, expressed as percentage lactic acid (w/v). Reducing sugars were quantified using the 3,5-dinitrosalicylic acid colorimetric method 13. Briefly, 1 mL of sample was mixed with 1 mL of DNS reagent, heated at 100°C for 5 min, cooled, and diluted to 10 mL, with absorbance measured at 540 nm.
2.4. HPLC Analysis of Organic Acids and SugarsSamples were filtered through 0.22 μm syringe filters and analyzed using a Shimadzu LC-20A high-performance liquid chromatography (HPLC) system (Shimadzu, Kyoto, Japan). Organic acids were separated on a Shodex RSpack KC-811 column (300 × 8 mm, Showa Denko, Tokyo, Japan) with a Shodex KC-G6B guard column, using 3.0 mmol/L perchloric acid as the mobile phase at 1.0 mL/min, 50°C, and detected at 210 nm (20 min run time). Sugars were resolved on a Shodex Sugar KS-801 column (300 × 8 mm, Showa Denko, Tokyo, Japan) with a KS-G6B guard column, using ultrapure water as the mobile phase at 0.7 mL/min, 80°C, with a 10 μL injection volume and refractive index detection (20 min run time).
2.5. Total Phenolic Content (TPC)Total phenolic content (TPC) was determined using the Folin-Ciocalteu method. Briefly, a 50 μL aliquot of ten-fold diluted sample was mixed with 50 μL of diluted Folin-Ciocalteu reagent, incubated at 25°C for 5 min, followed by the addition of 50 μL of 100 g/L Na2CO3 and adjustment to 250 μL with ultrapure water. After 60 min in the dark, absorbance was measured at 765 nm using a spectrophotometer. TPC was expressed as milligrams of gallic acid equivalents (GAE) per gram of fresh weight (FW).
2.6. Total Flavonoid Content (TFC)Total flavonoid content (TFC) was measured using the AlCl3 colorimetric method. A 50 μL sample was mixed with 25 μL of 50 g/L NaNO2, incubated for 5 min, followed by the addition of 25 μL of 100 g/L AlCl3 and a further 5 min incubation in the dark. Subsequently, 100 μL of 50 g/L NaOH was added, and the volume was adjusted to 250 μL with ultrapure water. Absorbance was recorded at 510 nm. TFC was expressed as milligrams of rutin equivalents (RE) per gram of fresh weight (FW).
2.7. Statistical AnalysisAll experiments were performed in triplicate, with results expressed as mean ± standard deviation (SD). Data were analyzed using SPSS 23.0 (IBM Corp., Armonk, NY, USA). The Mantel test was used to assess correlations between microbial growth and biochemical parameters, while principal component analysis (PCA) was applied to evaluate variations in biochemical profiles across strains and time points. One-way analysis of variance (ANOVA) with Tukey’s post-hoc test was conducted to identify significant differences (p < 0.05).
This study evaluated the fermentation of black tea-sugar broth by eight L. plantarum strains (HD2, JF2, J10, RU1, SC1, J30, GA6, LB3) over 48 h, focusing on microbial growth dynamics and their impact on biochemical parameters, including pH, titratable acidity, organic acids, reducing sugars, total phenolics, and flavonoids. The findings elucidate strain-specific metabolic profiles and their potential for developing functional fermented beverages.
3.1. Growth DynamicsThe growth profiles of the eight L. plantarum strains in black tea-sugar broth, assessed via viable cell counts (CFU/mL) at 0, 12, 24, 36, and 48 h, revealed distinct strain-specific patterns (Figure 1). All strains, inoculated at 3.0 × 106 CFU/mL, exhibited rapid proliferation within 12 h, entering the logarithmic growth phase, consistent with the adaptability of L. plantarum to tea-based substrates 14.
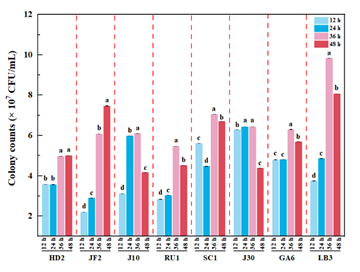
Among the strains, strain JF2 demonstrated sustained growth, increasing from 2.21 × 107 CFU/mL at 12 h to 7.47 × 107 CFU/mL at 48 h (p < 0.05), reflecting robust metabolic activity and environmental tolerance. This continuous growth suggests JF2’s suitability for extended fermentation processes, such as probiotic beverage production. In contrast, strain HD2 exhibited delayed growth, with viable counts remaining stable from 3.59 × 107 CFU/mL at 12 h to 3.57 × 107 CFU/mL at 24 h, before increasing to 5.01 × 107 CFU/mL at 48 h (p < 0.05). This extended lag phase may result from slower substrate utilization, as observed in other lactic acid bacteria fermentations 15.
Strains LB3, J10, J30, GA6, and RU1 reached peak viable counts at 36 h (9.84 × 107, 6.09 × 107, 6.44 × 107, 6.30 × 107, and 5.48 × 107 CFU/mL, respectively), followed by declines at 48 h (8.07 × 107, 4.17 × 107, 4.38 × 107, 5.69 × 107, and 4.52 × 107 CFU/mL, representing decreases of 18.0%, 31.5%, 32.0%, 9.7%, and 17.5%, respectively; p < 0.05). Such declines are typical in fermentation due to nutrient depletion or accumulation of inhibitory metabolites 16. LB3’s high peak and moderate decline suggest sustained viability, while J10 and J30’s sharper declines indicate lower tolerance to environmental stress.
Strain SC1 displayed a unique non-monotonic trend, decreasing from 5.61 × 107 CFU/mL at 12 h to 4.47 × 107 CFU/mL at 24 h, then increasing to 7.05 × 107 CFU/mL at 36 h, and slightly declining to 6.71 × 107 CFU/mL at 48 h (4.8% decrease). This pattern might reflect an initial adaptation phase followed by vigorous growth, possibly due to specific interactions with tea polyphenols or sugars. The observed variability in growth dynamics highlights strain-specific differences in proliferation rates and adaptation to the black tea-sugar broth matrix. These differences may influence the suitability of each strain for specific applications in tea-sugar broth fermentation.
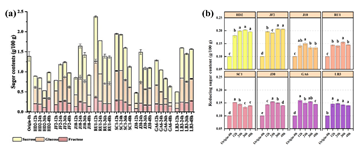
3.2. pH and Acid ChangesFermentation of black tea-sugar broth by L. plantarum strains resulted in significant acidification, characterized by decreased pH and increased titratable acidity (TA) (Figure 2a & Figure 2b). All strains reduced pH from 4.0–4.1 at 12 h to 3.78–3.98 at 48 h (p < 0.05), reflecting the accumulation of organic acids, particularly lactic and formic acids (Figure 2c). This acidification closely correlated with microbial growth dynamics, influencing the flavor, preservation, and probiotic functionality of the fermented products 17.

Strain JF2, exhibiting sustained growth (from 2.21 × 107 to 7.47 × 107 CFU/mL), reduced pH from 3.94 at 12 h to 3.78 at 48 h (p < 0.05), indicating strong acidogenic capacity. Similarly, strain HD2, despite delayed growth, lowered pH from 4.05 to 3.83 (p < 0.05), suggesting stable acid production post-lag phase. Strain LB3, peaking at 9.84 × 107 CFU/mL at 36 h, decreased pH from 4.06 to 3.88 (p < 0.05), consistent with its high viability. In contrast, strain SC1’s pH stabilized at 3.89 from 24 to 48 h, possibly due to buffering by tea compounds or reduced acid synthesis, despite its non-monotonic growth pattern. Strains J10 and J30, with significant growth declines after 36 h, showed moderate pH reductions (J10 from 4.03 to 3.87; J30 from 4.10 to 3.97; p < 0.05), suggesting acid accumulation may have limited further proliferation. Strains GA6 and RU1 exhibited milder pH decreases (from 4.06 to 3.95 and 4.07 to 3.98, respectively), aligning with their intermediate growth stability.
In the present study, TA levels were generally increased during fermentation. For instance, TA for strain HD2 rose from 3.20 mg/100 mL at 12 h to 3.43 mg/100 mL at 48 h, while that of strain LB3 increased from 2.25 mg/100 mL to 3.79 mg/100 mL, corresponding to their sustained or peak growth. However, strains JF2 and J10 showed slight TA declines after 24 h (JF2 from 2.96 to 2.61 mg/100 mL; J10 from 2.72 to 2.61 mg/100 mL), possibly due to acid reassimilation or metabolic conversion 18.
Lactic acid, the primary fermentation product via homofermentative glycolysis, dominated acidogenesis. Strain HD2’s lactic acid increased from 0.70 to 3.41 mg/100 mL (4.9-fold, p < 0.05), JF2 from 0.78 to 3.11 mg/100 mL (4.0-fold), and J10 from 1.39 to 3.22 mg/100 mL (2.3-fold), correlating with pH declines and growth. Strain LB3’s lactic acid rose modestly from 3.34 to 3.94 mg/100 mL (1.2-fold), reflecting early acidogenic activity consistent with its peak growth at 36 h. Strains SC1 and J30 showed increases (1.54 to 2.89 mg/100 mL and 0.76 to 3.57 mg/100 mL, respectively), moderated by pH dynamics.
Formic acid accumulation further contributed to acidification. Strain J10 showed the highest increase (from 1.68 to 7.62 mg/100 mL, 4.5-fold, p < 0.05), followed by HD2 (from 1.57 to 6.50 mg/100 mL, 4.1-fold) and JF2 (from 0.78 to 6.26 mg/100 mL, 8.0-fold). These increases may reflect activation of heterofermentative pathways under nutrient limitation or environmental stress 19. Strain LB3’s formic acid increased from 3.89 to 4.63 mg/100 g, complementing its lactic acid production.
In contrast, acetic, citric, and malic acids showed smaller but statistically significant changes (p < 0.05). For example, HD2’s acetic acid increased from 2.88 to 3.24 mg/100 mL (12.5%), citric acid from 0.38 to 0.42 mg/100 mL (10.5%), and malic acid from 0.50 to 0.62 mg/100 mL (24.0%). Despite their significance, their absolute increments were substantially lower than those of lactic acid (2.71 mg/100 mL) and formic acid (4.93 mg/100 mL), suggesting a limited contribution to pH reduction, likely due to their low initial concentrations and weak buffering capacity in the broth.
These acidogenic profiles highlighted strain-specific metabolic capacities and their impact on growth and fermentation outcomes. Rapidly growing strains like JF2 and LB3 produced substantial lactic and formic acids, driving pronounced pH declines, making them suitable for rapid fermentation processes. SC1’s stabilized pH and moderate acid production suggest suitability for products requiring controlled acidity. The growth declines of J10 and J30, despite significant acid production, indicate sensitivity to acid stress, underscoring the need for optimized conditions (e.g., pH buffering) to enhance viability. These findings inform strain selection for functional beverages, balancing growth efficiency and biochemical outcomes.
3.3. Changes in SugarDuring the fermentation of black tea-sugar broth by L. plantarum strains, sucrose, the primary carbon source, was hydrolyzed into glucose and fructose to support microbial growth and acid production. Over 48 h, sucrose concentrations displayed strain-dependent trends (Figure 3a), reflecting differences in invertase activity and sugar utilization efficiency. For example, in strain HD2, sucrose decreased from 0.28 g/100 g at 12 h to 0.19 g/100 g at 36 h, followed by a slight rebound to 0.25 g/100 g at 48 h, likely due to residual hydrolysis or delayed substrate consumption. In contrast, strain JF2 showed an initial increase from 0.28 to 0.43 g/100 g by 24 h, then declined to 0.37 g/100 g at 48 h, suggesting dynamic sucrose cleavage and potential re-assimilation. Strain SC1 exhibited a marked reduction from 0.96 to 0.53 g/100 g, indicating high invertase activity or greater initial sucrose availability 20. RU1 showed a similar trend, with sucrose declining from 1.13 to 0.65 g/100 g.
The hydrolysis products, glucose and fructose, accumulated transiently, peaking between 24 and 36 h, corresponding to the active fermentation phase. In strain JF2, glucose increased from 0.38 g/100 g at 12 h to 0.62 g/100 g at 36 h (p < 0.05), and fructose rose from 0.18 g/100 g to 0.26 g/100 g (p < 0.05), indicating efficient sucrose breakdown. Conversely, in strain HD2, glucose decreased from 0.43 g/100 g at 12 h to 0.24 g/100 g at 36 h, recovering to 0.42 g/100 g at 48 h, while fructose dropped from 0.19 g/100 g to 0.10 g/100 g before rising to 0.30 g/100 g. These fluctuations suggest a dynamic balance between monosaccharide accumulation and microbial consumption. Consistently, total reducing sugars (Figure 3b) peaked at 36 h (0.20 g/100 g for HD2; 0.21 g/100 g for JF2), followed by minor declines as monosaccharides were metabolized.
Moreover, strains J10 and J30 exhibited stable reducing sugar levels after 36 h (J10: 0.13 g/100 g; J30: 0.15 to 0.14 g/100 g), but marked declines in glucose and fructose were observed in J10, glucose from 0.58 to 0.30 g/100 g (−48.3%, p < 0.05) and fructose from 0.22 to 0.13 g/100 g (−40.9%, p < 0.05). This was accompanied by a drop in viable cell counts (6.09 × 107 to 4.17 × 107 CFU/mL), suggesting selective sugar depletion and stress-induced metabolic inhibition. High levels of formic acid (7.62 mg/100 g) and lactic acid (3.22 mg/100 g), along with a low pH of 3.87, likely exceeded the acid tolerance threshold of strain J10, impairing its growth and sugar utilization capacity 21. A similar pattern was observed in strain J30. In contrast, strains SC1 and RU1 exhibited relatively stable glucose and fructose profiles, despite slower sucrose degradation, implying a more conservative metabolic strategy, potentially due to lower invertase expression, warranting further enzymatic characterization.
Collectively, these results highlight a strong link between sucrose hydrolysis, monosaccharide dynamics, microbial proliferation, and acidogenesis. Rapid sucrose consumption between 12–36 h fueled lactic acid production via the Embden–Meyerhof–Parnas pathway 22. For strains HD2 and JF2, changes in sucrose levels (from 0.28 to 0.25 g/100 g for HD2; from 0.28 to 0.37 g/100 g for JF2) were well aligned with increases in lactic acid (HD2: from 0.70 to 3.41 mg/100 g; JF2: from 0.78 to 3.11 mg/100 g) and cell densities (HD2: from 3.59 × 107 to 5.01 × 107 CFU/mL; JF2: from 2.21 × 107 to 7.47 × 107 CFU/mL). In contrast, strains like J10 and J30, which experienced acid stress and lower sugar availability, demonstrated impaired growth in the later fermentation stages. These insights into sugar metabolism provide a metabolic basis for selecting suitable L. plantarum strains in functional beverage fermentation, balancing acid production and microbial viability.
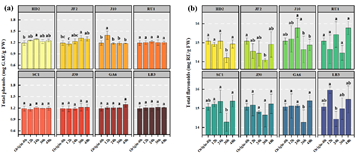
The contents of TPC and TFC are critical for the health benefits of fermented black tea-sugar broth, including antioxidant and anti-inflammatory properties 23. Their dynamics were monitored over 48 h to assess stability and interactions with microbial growth, acid production), and sugar metabolism (Figure 4).
As shown, the values of TPC and TFC remained stable across all strains, with variations typically below 15%. For stain HD2, TPC values ranged from 1.26 mg GAE/g FW at 12 h to 1.25 mg GAE/g FW at 48 h, and TFC values ranged from 14.92 to 14.93 mg RE/g FW (<1% change). Strain JF2 showed a slight increase in TPC values (from 1.16 to 1.30 mg GAE/g FW, 11.2%, p < 0.05) and TFC values (from 14.56 to 14.91 mg RE/g FW). In strain J10, it exhibited a minor decrease in TPC levels (from 1.39 to 1.18 mg GAE/g FW, 15.1%), while strain SC1’s TPC levels increased slightly (1.16 to 1.20 mg GAE/g FW, 3.4%). Strain RU1’s TFC values rose from 14.66 to 15.78 mg RE/g FW (7.6%). These minimal changes indicate that L. plantarum fermentation does not significantly degrade or synthesize these compounds, likely due to their antioxidant stability or the absence of degrading enzymes 24.
Stability was maintained despite metabolic changes. For strain HD2, stable values of TPC (1.23–1.29 mg GAE/g FW) and TFC (14.2–15.1 mg RE/g FW) persisted despite a pH drop (from 4.05 to 3.83), lactic acid increase (from 0.70 to 3.41 mg/100 g), and sucrose consumption (from 0.28 to 0.25 g/100 g). In strain JF2, the slight TPC increase coincided with robust growth (from 2.21 × 107 to 7.47 × 107 CFU/mL) and lactic acid production (from 0.78 to 3.11 mg/100 g), possibly due to minor release of bound phenolics. Strain J10’s TPC decrease aligned with reduced growth (6.09 × 107 to 4.17 × 107 CFU/mL) and sugar metabolism (glucose decreased from 0.58 to 0.30 g/100 g), suggesting limited phenolic extraction under stress. Strains LB3 and GA6 showed minor TFC changes (LB3: from 15.96 to 15.48 mg RE/g FW; GA6: from 15.13 to 15.39 mg RE/g FW), reinforcing stability during fermentation.
This stability is crucial for functional beverage development, as polyphenols and flavonoids contribute to antioxidant and cardiovascular benefits 25. The lack of significant degradation ensures retention of health attributes despite pH declines and acid/sugar metabolism, supporting sensory properties like astringency. The stability of these compounds also supports potential flavor retention, as polyphenols contribute to astringency and overall sensory appeal 26.
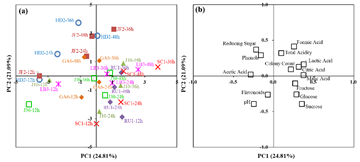
3.5. PCA ResultsTo elucidate the relationships among microbial growth, biochemical parameters, and fermentation time during the fermentation of black tea-sugar broth by L. plantarum strains, PCA was performed using the complete dataset. The analysis included eight strains (HD2, JF2, J10, RU1, SC1, J30, GA6, LB3) across four time points (12 h, 24 h, 36 h, 48 h). The first five principal components (PCs), with eigenvalues greater than 1, collectively accounted for 77.32% of the total variance, indicating that the model effectively captured the major patterns in the data. The first two PCs (PC1 and PC2), derived from the average values of the variables, accounted for 45.90% of the total variance: PC1 explained 24.81% and PC2 explained 21.09% (Figure 5).
The PCA score plot for PC1 versus PC2 is presented in Figure 5a. The samples exhibited distinct clustering based on fermentation time and strain-specific behaviors. PC1 primarily separated early fermentation stages (12 h and 24 h) from later stages (36 h and 48 h). For instance, strains J10 and LB3 fermented at 12 h (J10-12h and LB3-12h) were located in the negative PC1 region, while those fermented at 48 h (J10-48h and LB3-48h) shifted toward the positive PC1 quadrant, reflecting significant changes in biochemical profiles over time. PC2 highlighted strain-specific differences, with SC1 and RU1 samples showing greater dispersion, suggesting unique metabolic patterns.
The correlation loadings plot for PC1 and PC2 is shown in Figure 5b, revealing the contributions of individual variables to the PCs. PC1 was strongly negatively correlated with pH and positively correlated with total acidity, lactic acid, and formic acid, underscoring the dominant role of acidification in later fermentation stages. PC2 showed positive correlations with reducing sugars and colony counts, emphasizing the influence of sugar metabolism and microbial proliferation. For example, the positive PC1 shift of HD2-48h and JF2-48h aligned with increased lactic acid (HD2: 3.41 mg/100 g; JF2: 3.11 mg/100 g) and formic acid (HD2: 6.50 mg/100 g; JF2: 6.26 mg/100 g), confirming that acid production drove temporal variation. The dispersed distribution of strain SC1, with non-monotonic growth (5.61 × 107 to 6.71 × 107 CFU/mL) and stable pH (3.89), was consistent with the negative PC1 loading of pH. Moreover, strains J10 and J30 over 36 h located on negative PC2, correlating with sugar depletion (J10 glucose: 0.30 g/100 g) and declining colony counts (J10: 4.17 × 107 CFU/mL), supported by the positive PC2 loadings of reducing sugars and colony counts, which suggest sugar limitation and acid stress as growth constraints.
Overall, PCA effectively discriminated temporal and strain-specific effects, revealing complex interactions among sugar metabolism, acidification, and microbial growth. These findings provide valuable insights for optimizing fermentation processes in the development of functional fermented beverages. These PCA insights can guide industry in strain selection for tailored beverage profiles.
The fermentation of black tea-sugar broth by L. plantarum strains significantly influences microbial growth, acid production, sugar metabolism, and the stability of functional components, which are critical for the development of functional beverages. This study demonstrated that strain-specific metabolic profiles, including lactic and formic acid accumulation, sucrose hydrolysis, and stable total phenols and flavonoids, directly impact fermentation outcomes. Principal component analysis, explaining 77.32% of the variance, confirmed that acidification (PC1) and sugar metabolism with microbial proliferation (PC2) are key drivers of temporal and strain-specific variations. These findings indicate that strains like JF2 and LB3, with robust growth and acid production, are promising for rapid fermentation, while SC1’s stable pH suggests suitability for controlled acidity products. The stability of bioactive compounds, despite pH declines and sugar consumption, ensures retention of health benefits, such as antioxidant properties.
These results offer valuable insights for optimizing fermentation processes to enhance the functionality of black tea-based beverages, a growing area of interest in food science. However, the study highlights a critical gap in understanding the flavor profiles, as the aroma-producing capabilities of the eight L. plantarum strains remain unexplored. Future research should focus on elucidating specific metabolic pathways, such as invertase activity and phenolic stability mechanisms, while investigating the strains’ potential for aroma compound production using techniques like GC-MS, electronic nose, and electronic tongue. Additionally, assessing post-fermentation microbial viability and shelf life under storage conditions will be essential for commercial viability, given L. plantarum’s known acid tolerance. For industry adoption, our findings can inform kombucha or probiotic drink producers in selecting optimal strains, such as JF2 for balanced acidity and bioactive retention, potentially leading to innovative products with enhanced gut health benefits, extended shelf life, and greater consumer appeal.
The authors gratefully acknowledge the financial support from the National Natural Science Foundation of China (Nos. 32302267, 32302953).
| [1] | Guo, Q., Yuan, J., Ding, S., Nie, Q., Xu, Q., Pang, Y., Liao, X., Liu, Z., Liu, Z. and Cai. S. 2024. Microbial fermentation in fermented tea beverages: Transforming flavor and enhancing bioactivity. Beverage Plant Research. 4: e029. | ||
| In article | View Article | ||
| [2] | Frolova, Y., Vorobyeva, V., Vorobyeva, I., Sarkisyan, V., Malinkin, A., Isakov, V. and Kochetkova. A. 2023. Development of fermented kombucha tea beverage enriched with inulin and B vitamins. Fermentation. 9(6): 552. | ||
| In article | View Article | ||
| [3] | Jakubczyk, K., Łopusiewicz, Ł., Kika, J., Janda-Milczarek, K. and Skonieczna-Żydecka. K. 2024. Fermented tea as a food with functional value–its microbiological profile, antioxidant potential and phytochemical composition. Foods. 13(1): 50. | ||
| In article | View Article PubMed | ||
| [4] | Soumya, M.P. and Nampoothiri, K.M. 2021. An overview of functional genomics and relevance of glycosyltransferases in exopolysaccharide production by lactic acid bacteria. International Journal of Biological Macromolecules. 184: 1014–1025. | ||
| In article | View Article PubMed | ||
| [5] | Wang, Y., Wu, J., Lv, M., Shao, Zhen., Hungwe, M., Wang, J., Bai, X., Xie, J., Wang, Y. and Geng, W. 2021. Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Fronters in Bioengineering and Biotechnology. 9: 612285. | ||
| In article | View Article PubMed | ||
| [6] | Peng, Y., He, H., Li, Y., Zhao, J., Wang, B., Wei, Y., Yu, J., Chen, Q. and Wang, D. 2024. Fermentation of enzymatical feijoa fruit juice by probiotic bacteria: nutritional composition, bioactive phenolics, and antioxidant activities. Journal of Food Measurement and Characterization. 18:8564–8576. | ||
| In article | View Article | ||
| [7] | Guo, J., Wang, C., Liu, L., Xu, K., Li, B., Zhang, L., Xu, H. and Lei, H. 2023. Antioxidant activities and volatile compounds of Chinese cabbage sauce prepared by the combination of Lactobacillus plantarum and functional oligosaccharides. Food Bioscience. 54:102854. | ||
| In article | View Article | ||
| [8] | Zhou, Q., Xue, B., Gu, R., Li, P. and Gu, Q. 2021. Lactobacillus plantarum ZJ316 attenuates Helicobacter pylori-induced gastritis in C57BL/6 mice. Journal of Agricultural and Food Chemistry. 69(23): 6510–6523. | ||
| In article | View Article PubMed | ||
| [9] | Di Cagno, R., De Angelis, M., Limitone, A., Minervini, F., Carnevali, P., Corsetti, A., Gaenzle, M., Ciati, R. and Gobbetti, M. 2006. Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. Journal of Agricultural and Food Chemistry. 54(26): 9873–9881. | ||
| In article | View Article PubMed | ||
| [10] | Zhang, L., Liu, C., Li, D., Zhao, Y., Zhang, X., Zeng, X., Yang, Z. and Li. S. 2013. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. International Journal of Biological Macromolecules. 54: 270–275. | ||
| In article | View Article PubMed | ||
| [11] | Wang, S., Li, C., Xu, Q., Wang, Y., Wang, S., Zou, Y., Yang, Z. and Yuan, L. 2024. Addition of lactic acid bacteria modulates microbial community and promotes the flavor profiles of Kombucha. Food Bioscience. 60: 104340. | ||
| In article | View Article | ||
| [12] | Hassler, V., Brand, N. and Wefers, D. 2024. Isolation and characterization of exopolysaccharides from kombucha samples of different origins. International Journal of Biological Macromolecules. 267: 131377. | ||
| In article | View Article PubMed | ||
| [13] | Teixeira, G.G. and Santos, P.M. 2022. Simple and cost-effective approaches for quantification of reducing sugar exploiting digital image analysis. Journal of Food Composition and Analysis. 113: 104719. | ||
| In article | View Article | ||
| [14] | Lin, M., Lin, S., He, H., Yu, Y., Hu, J. and Zhou, L. 2025. Lactiplantibacillus plantarum in fermented beverages: Properties, mechanisms, and future prospects. Journal of Functional Foods. 129: 106864. | ||
| In article | View Article | ||
| [15] | Sharma, V. and Mishra H.N. 2014 Unstructured kinetic modeling of growth and lactic acid production by Lactobacillus plantarum NCDC 414 during fermentation of vegetable juices. LWT-Food Science and Technology. 59(2): 1123–1128. | ||
| In article | View Article | ||
| [16] | Gupta, S., Abu-Ghannam, N. and Scannell, A.G.M. 2011. Growth and kinetics of Lactobacillus plantarum in the fermentation of edible Irish brown seaweeds. Food and Bioproducts Processing. 89(4): 346–355. | ||
| In article | View Article | ||
| [17] | Aghababaie, M., Khanahmadi, M. and Beheshti, M. 2015. Developing a detailed kinetic model for the production of yogurt starter bacteria in single strain cultures. Food and Bioproducts Processing. 94: 657–667. | ||
| In article | View Article | ||
| [18] | Behera, S.S., Ray, R.C. and Zdolec, N. 2018. Lactobacillus plantarum with functional properties: An approach to increase safety and shelf-life of fermented foods. Biomed Research International. 2018: 9361614. | ||
| In article | View Article PubMed | ||
| [19] | Garcia-Gonzalez, N., Battista, N., Prete, R. and Corsetti, A. 2021. Health-promoting role of Lactiplantibacillus plantarum isolated from fermented foods. Microorganisms. 9(2): 349. | ||
| In article | View Article PubMed | ||
| [20] | Cohen, G., Sela, D.A. and Nolden, A.A. 2023. Sucrose concentration and fermentation temperature impact the sensory characteristics and liking of Kombucha. Foods. 12(16): 3116. | ||
| In article | View Article PubMed | ||
| [21] | Derunets, A.S., Selimzyanova, A.I., Rykov, S.V., Kuznetsov, A.E. and Berezina, O.V. 2024. Strategies to enhance stress tolerance in lactic acid bacteria across diverse stress conditions. World Journal of Microbiology and Biotechnology. 40: 126. | ||
| In article | View Article PubMed | ||
| [22] | Wen, L., Yang, L., Chen, C., Li, J., Fu, J., Liu, G., Kan, Q., Ho, C.T., Huang, Q., Lan, Y. and Cao, Y. 2024. Applications of multi-omics techniques to unravel the fermentation process and the flavor formation mechanism in fermented foods. Critical Reviews in Food Science and Nutrition. 64(23): 8367–8383. | ||
| In article | View Article PubMed | ||
| [23] | Shi, S., Wei, Y., Lin, X., Liang, H., Zhang, S., Chen, Y., Dong, Li. and Ji, C. 2023. Microbial metabolic transformation and antioxidant activity evaluation of polyphenols in kombucha. Food Bioscience. 51: 102287. | ||
| In article | View Article | ||
| [24] | Romero, M.L.M., Staszewski, M. and Martínez, M.J. 2021. The effect of green tea polyphenols addition on the physicochemical, microbiological and bioactive characteristics of yogurt. British Food Journal. 123(7): 2380–2397. | ||
| In article | View Article | ||
| [25] | Wu, S.X., Xiong, R.G., Cheng, J., Xu, X.Y., Tang, G.Y., Huang, S.Y., Zhou, D.D., Saimaiti, A., Gan, R.Y. and Li, H.B. 2023. Preparation, antioxidant activities and bioactive components of kombucha beverages from golden-flower tea (Camellia petelotii) and honeysuckle-flower tea (Lonicera japonica). Foods. 12(16): 3010. | ||
| In article | View Article PubMed | ||
| [26] | Xing, Z., Fu, X., Huang, H., Xu, Y., Wei, L., Shan, C. and Du, Y. 2025. Recent advances in Lactobacillus plantarum fermentation in modifying fruit-based products: Flavor property, bioactivity, and practical production applications. Comprehensive Reviews in Food Science and Food Safety. 24(2): e70160. | ||
| In article | View Article PubMed | ||
Published with license by Science and Education Publishing, Copyright © 2025 Zhiyuan Wang, Jiayi Yu, Yu Su, Jiaxiu Wang, Shuai Wang, Lei Yuan and Shuo Wang
![]() This work is licensed under a Creative Commons Attribution 4.0 International License. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/
This work is licensed under a Creative Commons Attribution 4.0 International License. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/
| [1] | Guo, Q., Yuan, J., Ding, S., Nie, Q., Xu, Q., Pang, Y., Liao, X., Liu, Z., Liu, Z. and Cai. S. 2024. Microbial fermentation in fermented tea beverages: Transforming flavor and enhancing bioactivity. Beverage Plant Research. 4: e029. | ||
| In article | View Article | ||
| [2] | Frolova, Y., Vorobyeva, V., Vorobyeva, I., Sarkisyan, V., Malinkin, A., Isakov, V. and Kochetkova. A. 2023. Development of fermented kombucha tea beverage enriched with inulin and B vitamins. Fermentation. 9(6): 552. | ||
| In article | View Article | ||
| [3] | Jakubczyk, K., Łopusiewicz, Ł., Kika, J., Janda-Milczarek, K. and Skonieczna-Żydecka. K. 2024. Fermented tea as a food with functional value–its microbiological profile, antioxidant potential and phytochemical composition. Foods. 13(1): 50. | ||
| In article | View Article PubMed | ||
| [4] | Soumya, M.P. and Nampoothiri, K.M. 2021. An overview of functional genomics and relevance of glycosyltransferases in exopolysaccharide production by lactic acid bacteria. International Journal of Biological Macromolecules. 184: 1014–1025. | ||
| In article | View Article PubMed | ||
| [5] | Wang, Y., Wu, J., Lv, M., Shao, Zhen., Hungwe, M., Wang, J., Bai, X., Xie, J., Wang, Y. and Geng, W. 2021. Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Fronters in Bioengineering and Biotechnology. 9: 612285. | ||
| In article | View Article PubMed | ||
| [6] | Peng, Y., He, H., Li, Y., Zhao, J., Wang, B., Wei, Y., Yu, J., Chen, Q. and Wang, D. 2024. Fermentation of enzymatical feijoa fruit juice by probiotic bacteria: nutritional composition, bioactive phenolics, and antioxidant activities. Journal of Food Measurement and Characterization. 18:8564–8576. | ||
| In article | View Article | ||
| [7] | Guo, J., Wang, C., Liu, L., Xu, K., Li, B., Zhang, L., Xu, H. and Lei, H. 2023. Antioxidant activities and volatile compounds of Chinese cabbage sauce prepared by the combination of Lactobacillus plantarum and functional oligosaccharides. Food Bioscience. 54:102854. | ||
| In article | View Article | ||
| [8] | Zhou, Q., Xue, B., Gu, R., Li, P. and Gu, Q. 2021. Lactobacillus plantarum ZJ316 attenuates Helicobacter pylori-induced gastritis in C57BL/6 mice. Journal of Agricultural and Food Chemistry. 69(23): 6510–6523. | ||
| In article | View Article PubMed | ||
| [9] | Di Cagno, R., De Angelis, M., Limitone, A., Minervini, F., Carnevali, P., Corsetti, A., Gaenzle, M., Ciati, R. and Gobbetti, M. 2006. Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. Journal of Agricultural and Food Chemistry. 54(26): 9873–9881. | ||
| In article | View Article PubMed | ||
| [10] | Zhang, L., Liu, C., Li, D., Zhao, Y., Zhang, X., Zeng, X., Yang, Z. and Li. S. 2013. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. International Journal of Biological Macromolecules. 54: 270–275. | ||
| In article | View Article PubMed | ||
| [11] | Wang, S., Li, C., Xu, Q., Wang, Y., Wang, S., Zou, Y., Yang, Z. and Yuan, L. 2024. Addition of lactic acid bacteria modulates microbial community and promotes the flavor profiles of Kombucha. Food Bioscience. 60: 104340. | ||
| In article | View Article | ||
| [12] | Hassler, V., Brand, N. and Wefers, D. 2024. Isolation and characterization of exopolysaccharides from kombucha samples of different origins. International Journal of Biological Macromolecules. 267: 131377. | ||
| In article | View Article PubMed | ||
| [13] | Teixeira, G.G. and Santos, P.M. 2022. Simple and cost-effective approaches for quantification of reducing sugar exploiting digital image analysis. Journal of Food Composition and Analysis. 113: 104719. | ||
| In article | View Article | ||
| [14] | Lin, M., Lin, S., He, H., Yu, Y., Hu, J. and Zhou, L. 2025. Lactiplantibacillus plantarum in fermented beverages: Properties, mechanisms, and future prospects. Journal of Functional Foods. 129: 106864. | ||
| In article | View Article | ||
| [15] | Sharma, V. and Mishra H.N. 2014 Unstructured kinetic modeling of growth and lactic acid production by Lactobacillus plantarum NCDC 414 during fermentation of vegetable juices. LWT-Food Science and Technology. 59(2): 1123–1128. | ||
| In article | View Article | ||
| [16] | Gupta, S., Abu-Ghannam, N. and Scannell, A.G.M. 2011. Growth and kinetics of Lactobacillus plantarum in the fermentation of edible Irish brown seaweeds. Food and Bioproducts Processing. 89(4): 346–355. | ||
| In article | View Article | ||
| [17] | Aghababaie, M., Khanahmadi, M. and Beheshti, M. 2015. Developing a detailed kinetic model for the production of yogurt starter bacteria in single strain cultures. Food and Bioproducts Processing. 94: 657–667. | ||
| In article | View Article | ||
| [18] | Behera, S.S., Ray, R.C. and Zdolec, N. 2018. Lactobacillus plantarum with functional properties: An approach to increase safety and shelf-life of fermented foods. Biomed Research International. 2018: 9361614. | ||
| In article | View Article PubMed | ||
| [19] | Garcia-Gonzalez, N., Battista, N., Prete, R. and Corsetti, A. 2021. Health-promoting role of Lactiplantibacillus plantarum isolated from fermented foods. Microorganisms. 9(2): 349. | ||
| In article | View Article PubMed | ||
| [20] | Cohen, G., Sela, D.A. and Nolden, A.A. 2023. Sucrose concentration and fermentation temperature impact the sensory characteristics and liking of Kombucha. Foods. 12(16): 3116. | ||
| In article | View Article PubMed | ||
| [21] | Derunets, A.S., Selimzyanova, A.I., Rykov, S.V., Kuznetsov, A.E. and Berezina, O.V. 2024. Strategies to enhance stress tolerance in lactic acid bacteria across diverse stress conditions. World Journal of Microbiology and Biotechnology. 40: 126. | ||
| In article | View Article PubMed | ||
| [22] | Wen, L., Yang, L., Chen, C., Li, J., Fu, J., Liu, G., Kan, Q., Ho, C.T., Huang, Q., Lan, Y. and Cao, Y. 2024. Applications of multi-omics techniques to unravel the fermentation process and the flavor formation mechanism in fermented foods. Critical Reviews in Food Science and Nutrition. 64(23): 8367–8383. | ||
| In article | View Article PubMed | ||
| [23] | Shi, S., Wei, Y., Lin, X., Liang, H., Zhang, S., Chen, Y., Dong, Li. and Ji, C. 2023. Microbial metabolic transformation and antioxidant activity evaluation of polyphenols in kombucha. Food Bioscience. 51: 102287. | ||
| In article | View Article | ||
| [24] | Romero, M.L.M., Staszewski, M. and Martínez, M.J. 2021. The effect of green tea polyphenols addition on the physicochemical, microbiological and bioactive characteristics of yogurt. British Food Journal. 123(7): 2380–2397. | ||
| In article | View Article | ||
| [25] | Wu, S.X., Xiong, R.G., Cheng, J., Xu, X.Y., Tang, G.Y., Huang, S.Y., Zhou, D.D., Saimaiti, A., Gan, R.Y. and Li, H.B. 2023. Preparation, antioxidant activities and bioactive components of kombucha beverages from golden-flower tea (Camellia petelotii) and honeysuckle-flower tea (Lonicera japonica). Foods. 12(16): 3010. | ||
| In article | View Article PubMed | ||
| [26] | Xing, Z., Fu, X., Huang, H., Xu, Y., Wei, L., Shan, C. and Du, Y. 2025. Recent advances in Lactobacillus plantarum fermentation in modifying fruit-based products: Flavor property, bioactivity, and practical production applications. Comprehensive Reviews in Food Science and Food Safety. 24(2): e70160. | ||
| In article | View Article PubMed | ||



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}