Ectoine, a cyclic amino acid derivative is produced by some moderately halophilic microorganisms. It is widely used in the field of medicine and cosmetics; however its high cost, low yield and complicated production process restrict its use in an industry. The present study was carried out to construct an engineered Escherichia coli for the heterologous ectoine production. Firstly, the ectoine synthesis cluster (ectABC genes) from Halomonas elongata were introduced into E. coli for the ectoine production in heterologous system. Secondly, the effect of deletion for lysA and thrA on producing ectoine was investigated. Thirdly, the bioconversion conditions for synthesizing ectoine were optimized. In this study, nine strains of E.coli BW25113 were constructed, and an engineering strain ABC (∆lysA)/BW25113 with ectoine synthesis ability was screened out. An improved yield of ectoine was obtained with optimized conditions like 100 mmol/L sodium aspartate, 100 mmol/L glycerol, 80 mmol/L glucose and 120 mmol/L KCl at temperature 30°C, and the pH 7.0. A yield 2.08 g/L of ectoine was obtained. After preliminary optimization, the ABC (∆lysA)/BW25113 catalytic synthesis of ectoine increased by 2 times, indicating that optimized reaction conditions can increase the yield of ectoine. Here, we successfully constructed an engineered Escherichia coli for the heterologous production of ectoine. Our study herein provides a feasible and valuable biosynthesis pathway of ectoine with a potential for large-scale industrial production using simple and cheap feedstocks.
1, 4, 5, 6-tetrahydro-2-methyl-4-pyrimidine carboxylic acid is also known as ectoine (Tetrahydropyrimidine). It is a kind of cyclic amino acid derivative that was first discovered in Ectothiorhodospira halochloris by Galinski et al 1. The synthetic pathway of ectoine exists in many aerobic heterotrophic bacteria, such as Rhodotorula, Halomonas, Halobacterium, Vibrio, r-protein bacteria and Pseudomonas, even archaea nitroso-algae or basophilic methoxigenes and so on. The natural synthesis pathway of ectoine is only in some moderately halophilic microorganisms 2. Ectoine is a polar, easily soluble, and uncharged small molecule organic substance in the physiological pH range. It can stabilize cell swelling pressure without hindering the normal physiological functions of cells 3, and can also be used as a molecular chaperone 4. As a high-quality enzyme, protein, and cell protectant, it has more extensive applications in the medical, cosmetic, and biological industries. The current trading price of ectoine is 1,000 USD/kg, which is a high-priced commodity on the market 5.
So far, the ectABC gene cluster for ectoine biosynthesis has been found in many microorganisms 6. The synthesis pathway of ectoine belongs to the branch of aspartate metabolism. It starts from L-Aspartate 4-semialdehyde and enters a specific pathway 7. Under the catalysis of three enzymes encoded by the ectABC gene, Ectoine is synthesized through a three-step reaction. The ectB gene encodes L-2, 4-diaminobutyrate aminotransferase, which catalyzes the first step from L-Aspartate-4-semialdehyde to L-2, 4-diaminobutyrate; ectA gene encodes L-2, 4-aminobutyrate acetyltransferase, which catalyzes the second step of L-2, 4-diaminobutyrate to N-acetyl-2, 4-diaminobutyrate; ectC gene encodes ectoine synthase, which catalyzes the third step of N-acetyl-2, 4-diaminobutyrat cyclization to form ectoine. In addition, studies have shown that there is another ectoine synthesis pathway in a few microorganisms, which can use glutamate to catalyze the synthesis of ectoine, but the detailed reaction steps are not clear in current research 8. In some studies, another synthetic pathway that skips ectoine has been found. It is postulated that N-acetyl-2, 4-diaminobutyrate could be directly converted into 3-Hydroxy-N-acetyl-2.4-diaminobutyrate, and was then further converted into 5-hydroxytetrahydropyrimidine. However, this pathway is less reported and not very clear 9. As shown in Figure 1, in E. coli, L-aspartate-β-semialdehyde is the precursor for both L-lysine synthesis and ectoine synthesis 10. Thus, an adequate supply of L-aspartate-β-semialdehyde precursor for ectoine biosynthesis was expectedly obtained following the elimination of L-lysine synthesis pathway.
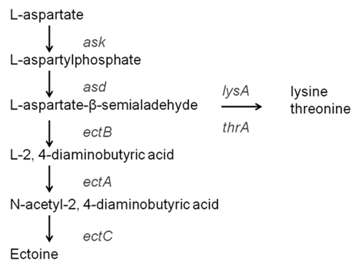
At present, the main production method of ectoine is microbial fermentation, which is divided into direct fermentation and whole-cell catalysis. The engineering strains commonly used for the production of ectoine by direct fermentation is halophilic bacteria 11, E. coli and Corynebacterium glutamicum, while the whole cell catalysis method mainly uses Escherichia coli 12. Compared with the direct fermentation method, the whole-cell catalysis method can avoid the problem of high salt induction caused by halophilic bacteria, does not corrode the fermenter and related equipments, and has the advantage of mild reaction conditions. Whole-cell catalysis uses E. coli, which is easier to cultivate and realize industrialization than other bacteria. In addition, the production of ectoine by whole-cell catalysis also has the advantages of less impurity content, easy separation and purification of products, and reusable bacteria. Therefore, ectoine synthesis by whole-cell catalysis has been highly regarded in recent years 13. This study intends to synthesize ectoine with high value by using low-priced and readily available sodium aspartate and glycerol as substrates. Using E. coli BW25113 as a host, an engineered strain for synthesizing ectoine was constructed. the whole-cell catalytic conditions of the engineered strain were optimized to obtain a strain with high ectoine production optimized conditions, which sets the foundation of industrialization of ectoine.
Chemicals for selection such as Ampicillin and Isopropyl 1-B-D-thiogalactopyranoside (IPTG), L-arabinose were brought from Takara Bio Ltd., (Hangzhou, China). Other chemicals were commercially obtained as an analytical grade. Acetonitrile (HPLC grade) and ectoine of 95% purity were of the highest quality and purchased from Sigma Aldrich (Shanghai, China). Media components such as tryptone, yeast extract and other chemicals were of analytical grade from Oxoid Co., Ltd. (Basingstoke, England). E. coli BL21 and DH5α competent cells were purchased from TSINGKE (Hangzhou China). All engineered E. coli strains was grown in Luria–Bertani (LB) consisting of 1.0% (w/v) tryptone, 0.5% (w/v) yeast extract, and 1% (w/v) NaCl. One step cloning kit was purchased from Vazyme (Nanjing, China).
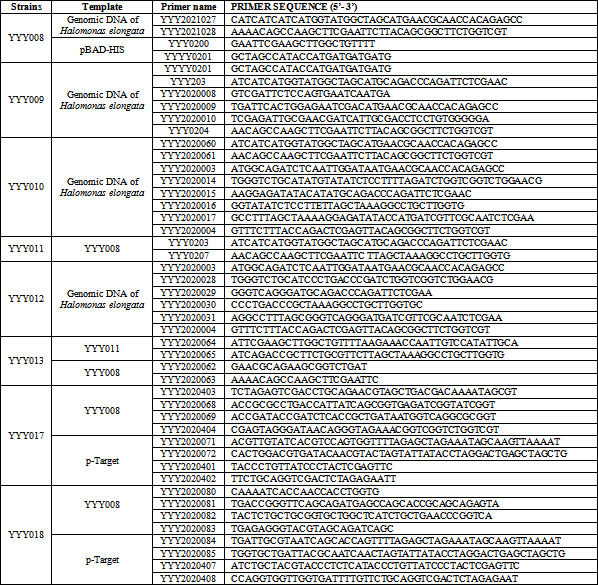
2.2. Construction of Recombinant Expression StrainsThe published ectoine biosynthetic cluster sequence of H. elongata in this study was used 14. The primers used in this study was presented in Table 1. The ectABC gene was amplified with ectABC-F and ectABC-R using pBAD/HisA-ectABC as the template, then subsequently cloned into pETDuet-1, resulting in pETDuet-ectABC. The recombinant plasmid pBAD/HisA-ectABC was transformed into BL21 (DE3) and BW25113, resulting in YYY001, and YYY008, respectively. The recombinant plasmid pETDuet-ectABC was transformed into BL21, resulting in YYY002. Deletion of lysA gene or thrA was performed using the CRISPR-Cas9 system according to the previously described method 15. Firstly, A 20 bp lysA gRNA or thrA gRNA nucleotides was introduced into the pTarget plasmid using primers YYY2020071 + YYY2020072, and YYY2020084 + YYY2020085, respectively, resulting in pTarget-lysA and pTarget-thrA.
The upstream and downstream homologous arms of lysA were amplified by YYY2020403 + YYY2020068 and YYY2020069 + YYY2020404 using genomic DNA BW25113 as the template. The upstream and downstream homologous arms of lysA was overlapped by overlap PCR, then subsequently cloned into pTarget-lysA using one step cloning kit, resulting in YYY017. Finally, the deletion strain for lysA was obtained by transforming YYY017 into cells.
The upstream and downstream homologous arms of thrA were amplified by YYY2020080 + YYY2020081, and YYY2020082 + YYY2020083 using genomic DNA BW25113 as the template. The upstream and downstream homologous arms of thrA were overlapped by overlap PCR, then subsequently cloned into pTarget-thrA using one step cloning kit, resulting in YYY018. Finally, the deletion strain for thrA was obtained by transforming YYY018 into cells.
The upstream and downstream homologous arms of thrA were amplified by YYY2020080 + YYY2020081, and YYY2020082 + YYY2020083 using genomic DNA BW25113 as the template. The upstream and downstream homologous arms of thrA were overlapped by overlap PCR, then subsequently cloned into pTarget-thrA using one step cloning kit, resulting in YYY018. Finally, the deletion strain for thrA was obtained by transforming YYY018 into cells.
The recombinant strains were grown at 37 °C in LB medium with ampicillin. When the optical density of the culture at 600 nm reached 0.6, 1 g/L L-arabinose or 0.1 mM IPTG was added into cells and incubated at 30 °C for 12 h.
2.4. Bioconversion ConditionsAfter induction, cells were collected by centrifugation at 4500rpm for 5 min, and then resuspended in the reaction mixture containing sterile water (pH 7.0), 50 mM sodium aspartate, 50 mM KCl, and 100 mM glycerol, 60 mM glucose to form a cell suspension (OD600 = 50). The optimization of bioconversion reaction conditions was performed using cell suspensions in 250 mL flasks in which the final liquid volume was 30 mL at 30°C and 200 rpm on a rotary shaker for 28 h. To optimize the component of reaction mixture, various concentrations of sodium aspartate (50, 100, 150, 200 and 250 mM) were compared in a bioconversion test. KCl has been reported to improve the activity and stability of ectB 16, so various concentrations of KCl (50, 80, 120, 150, and 200 mM) were added into the reaction mixture to investigate the effect on ectoine production. To evaluate the effect of glycerol on ectoine bioconversion, different concentrations of glycerol (20, 50, 80, 100 and 150 mM) were added into the reaction mixture. To evaluate the effect of glucose on ectoine bioconversion, different glucose concentrations (50, 60, 70, 80 and 90 mM) were added into the reaction mixture. To evaluate the effect of temperature on ectoine bioconversion, the reactions were performed at 30°C, at pH 7.0. To estimate the effect of pH, the reaction mixture was adjusted to pH levels of 5.5, 6.0, 6.5, 7.0, 7.5, and 8.0, and incubated at 30°C.
2.5. Cell Biomass MeasurementBiomass concentrations were estimated by measuring the optical density at 600 nm (OD600). The dry cell weight (DCW) was calculated based on OD600 (1 OD/mL =0.31 g DCW/L).
2.6. HPLC Analytical MethodsFor the identification of intracellular ectoine, cells were harvested by centrifugation. The samples were filtered through a 0.22-um pore size membrane filter and then analysed by isocratic HPLC using a Welchrom C18, 5μm, 4.6X 250mm column (4.6 × 250 mm, 5 um) with acetonitrile/water (2:98 v/v) at a flow rate of 1 mL/min as the mobile phase.
E. coli strains like like DH5α, BW25113 and BL21 were earlier used to synthesize ectoine 17, 18. To compare which hosts were more suitable for the synthesis of ectoine, the ectABC genes from H. elongata were amplified and expressed in BW25113 and BL21(DE3) under the control of T7 promoter, resulting in YYY001 and YYY002, as shown in Figure 2.
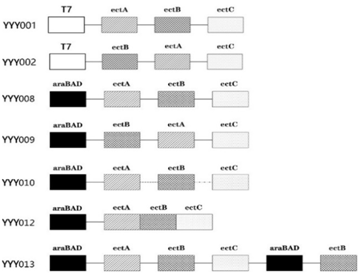
Both YYY001 and YYY002 were induced by the same concentration of IPTG (0.2 mM). As shown in Figure 3, the amount of ectoine synthesized by YYY001 was higher than YYY002. In order to compare the IPTG inducible T7 promoter with arabinose-inducible araBAD promoter used for synthesis of ectoine, the ectABC genes was cloned and expressed in BW25113, resulting in YYY008. As shown in Figure 3, the amount of ectoine synthesized by YYY008 was much higher than YYY0002 and YYY001. Therefore, in the further study BW25113 cells were choosen as the appropriate host cells, and araBAD promoter was chosen as the appropriate promoter for the synthesis of ectoine.
3.2. The Importance of ectABC Genes Order for Ectoine Biosynthesis in E. coliIt has been reported that engineering ribosome binding sites (RBS) strength are important for tuning multi-gene expression levels in E.coli 19. We constructed one strain named YYY010, in which the natural RBS inside ectABC cluster was changed into the RBS from commercial plasmid pBAD-HIS (AAGGAG). As shown in Figure 3, the amount of ectoine synthesized in YYY010 was lower than in YYY008. This indicated that the natural RBS is important for the efficient expression of ectABC genes.
EctC catalyzed the last step in ectoine synthesis through cyclo-condensation of the EctA-formed substrate N-gamma-acetyl-L-2, 4-diaminobutyric acid via a water elimination reaction 20. And the expression level of EctC was lower than EctA and EctB. The low level of ectC might restrict the efficiency of ectoine biosynthesis. To address this, the ectABC genes were fused in YYY012 with one special linker (GGGTCAGGG), which was reported to have high catalytic efficiency and bioactivities 21, 22. As shown in Figure 3, the amount of ectoine synthesized in YYY012 was lower than in YYY008. This indicated that this fusion protein might have a low catalytic activity due to partially un-functional caused by the stereo-hindrance effect.
It has been reported that EctB catalyzing the first step of ectoine biosynthesis is the rate-limiting enzyme involved in ectoine biosynthesis 23. The effect of ectABC order on ectoine synthesis was also investigated. The ectABC order was changed into ectBAC in YYY009. As shown in Figure 3, the amount of ectoine synthesized in YYY009 was lower than in YYY008. This indicated that the ectABC order is more efficient than ectBAC for ectoine biosynthesis.
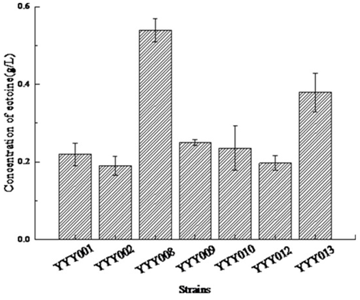
Since ectB is the rate-limiting enzyme, an additional copy of ectB was over-expressed with arabinose-inducible araBAD promoter in YYY013 (Figure 3). Interestingly, there is no any increase of ectoine was observed in this ectB over-expressed strain. This indicated that the ratio of ectA, ectB, ectC genes might be crucial for ectoine biosynthesis.
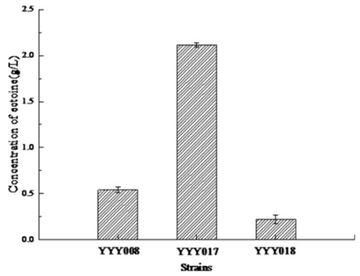
3.3. Deletion of lysA Improved the Yield of EctoineThe lysine pathway starts from l-Aspartate-4-semialdehyde, and the selection of lysA by gene knockout can reduce the carbon metabolism of this pathway, and remove the feedback inhibition of lysine on the aspartate kinase. Using Cas9/gRNA system, the lysA gene and thrA were deleted in YYY008, resulting in YYY017 (△lysA) and YYY018 (△thrA), respectively.
Interestingly, the amount of ectoine synthesized in YYY017 was higher than in YYY008, indicating that removing lysine can significantly increase the ectoine biosynthesis (Figure 4). However, deletion of thrA did not increase the yield of ectoine in YYY0017, which indicated that thrA might be a key gene involved in feedback inhibition of aspartate kinase (Figure 4).
The production of ectoine by the whole-cell catalytic method has the advantages of less impurity, easy separation and purification of the product, and reuse of the bacteria. To increase the yield of ectoine, the whole-cell catalytic system was optimized, the concentrations of sodium aspartate (substrate for the synthesis of ectoine), glycerol (substrate and energy for the production of acetyl-coa in EctA), KCl (Stabilizing EctB activity), temperature (affecting enzyme activity) and pH (affecting enzyme activity) were included.
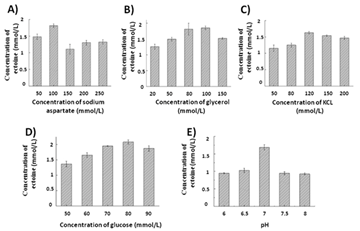
Sodium aspartate is the substrate for the ectoine synthesis, and its concentration is one of the factors affecting the yield of ectoine. Sodium aspartate is the substrate for the ectoine synthesis, and its concentration is one of the factors affecting the yield of ectoine. Therefore, the effects of different concentrations of sodium aspartate on the synthesis of ectoine were compared. As shown in Figure 5A, the yield of ectoine increased at first and then decreased with the increase of sodium aspartate concentration. When the concentration of sodium aspartic acid was 100 mmol/L, the yield of ectoine was the highest, 1.89 g/L. When the concentration of sodium aspartic acid was increased to 150 and 200 mmol/ L, the yield of ectoine did not increase further. When the concentration of sodium aspartic acid was 300 mmol /L, only 1.32 g/ L ectoine was generated. Therefore, the optimal concentration of sodium aspartic acid is 100 mmol/L.
Glycerol can provide substrate and energy for the production of acetyl-CoA in EctA catalysis and is one of the factors affecting the synthesis of ectoine. Therefore, the effect of glycerol on the synthesis of ectoine was further investigated. As shown in Figure 5B, as the concentration of glycerol increases, the yield of ectoine increases first and then stabilizes and then decreases. When the glycerol was 100 mmol/L, the yield of ectoine was the highest, about 1.85 g/L. When the glycerol concentration was further increased to 150 mmol/L, only 1.52 g/L of ectoine was produced. Therefore, the optimal glycerol concentration is 100 mmol/L.
KCl has the effect of stabilizing the activity of EctB. To understand the impact of KCl on the yield of ectoine, the effect of different concentrations of KCl on the synthesis of ectoine was investigated. As shown in Figure 5C, the yield of ectoine increased at first and then decreased with the increase of KCl concentration. When the concentration of KCl was 120 mmol/l, the yield of ectoine reached a maximum of 1.63 g/l. When there was a higher concentration of KCl (200 mmol/L), the yield of ectoine decreased significantly. Therefore, the optimal concentration of KCl was 120 mmol/L.
Glucose, a precursor of L-aspartic Acid, plays an important role in the synthesis of ectoine. To understand the effect of glucose on the yield of ectoine, the effects of different concentrations of glucose on the synthesis of ectoine were investigated. As shown in Figure 5D, the yield of ectoine increased at first and then decreased with the increase of KCl concentration. When the glucose concentration was 80 mmol/l, the yield of ectoine reached the maximum of 2.08 g/l. When the concentration of glucose was 90 mmol/l, the yield of ectoine decreased significantly. Therefore, the optimal concentration of glucose was 80 mmol/L.
PH is one of the important factors affecting enzyme activity, so the effect of pH on the synthesis of ectoine was investigated. As shown in Figure 5E, the yield of ectoine increased at first and then decreased with the increase of pH. When pH is 7.0, the yield of ectoine reaches a maximum of 1.8±0.5 g/L. However, when pH was higher than 7.5 and 8.0, the yield of ectoine was 0.95 g/L. Therefore, the optimal pH is 7.0.
In this study, an engineered strain of E.coli BW25113 namely ABC (∆lysA) /BW25113 showed improved ectoine synthesis ability. Optimized reaction conditions like 100 mmol/l sodium aspartate, 100 mmol/L glycerol, 80 mmol/L glucose and 120 mmol/L KCl at temperature 30°C, and the pH 7.0 were found to improve the yield of ectoine. However, there are still some problems to be solved and studied, such as the obvious differences in the expression of EctA, EctB, EctC, whether it is one of the important factors to limit the increase of ectoine yield, and the isolation and purification of ectoine have not been studied.
This study was financially supported by the Zhejiang Provincial Natural Science Foundation of China (Grant Nos. LQ18C010006 and LY19C010005), and National Natural Science Foundation of China (Grant Number: 31900497).
| [1] | Erwin A. GALINSKI’ H-PPaHGT: 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxy~c acid A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. 1985. | ||
| In article | View Article PubMed | ||
| [2] | Chen WC, Hsu CC, Lan JC, Chang YK, Wang LF, Wei YH: Production and characterization of ectoine using a moderately halophilic strain Halomonas salina BCRC17875. J Biosci Bioeng 2018, 125:578-84. | ||
| In article | View Article PubMed | ||
| [3] | Chen W, Zhang S, Jiang P, Yao J, He Y, Chen L, Gui X, Dong Z, Tang SY: Design of an ectoine-responsive AraC mutant and its application in metabolic engineering of ectoine biosynthesis. Metab Eng 2015, 30:149-55. | ||
| In article | View Article PubMed | ||
| [4] | Judith Becker1† RS, Michael Kohlstedt1, Björn J Harder1, Nicole S Borchert1, Nadine Stöveken2,3, Erhard Bremer2,3 and Christoph Wittmann1*: Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. 2013. | ||
| In article | |||
| [5] | Liu M, Liu H, Shi M, Jiang M, Li L, Zheng Y: Microbial production of ectoine and hydroxyectoine as high-value chemicals. Microb Cell Fact 2021, 20:76. | ||
| In article | View Article PubMed | ||
| [6] | Schwibbert K, Marin-Sanguino A, Bagyan I, Heidrich G, Lentzen G, Seitz H, Rampp M, Schuster SC, Klenk HP, Pfeiffer F, Oesterhelt D, Kunte HJ: A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581 T. Environ Microbiol 2011, 13:1973-94. | ||
| In article | View Article PubMed | ||
| [7] | Giesselmann G, Dietrich D, Jungmann L, Kohlstedt M, Jeon EJ, Yim SS, Sommer F, Zimmer D, Muhlhaus T, Schroda M, Jeong KJ, Becker J, Wittmann C: Metabolic Engineering of Corynebacterium glutamicum for High-Level Ectoine Production: Design, Combinatorial Assembly, and Implementation of a Transcriptionally Balanced Heterologous Ectoine Pathway. Biotechnol J 2019, 14:e1800417. | ||
| In article | View Article PubMed | ||
| [8] | Pastor JM, Salvador M, Argandona M, Bernal V, Reina-Bueno M, Csonka LN, Iborra JL, Vargas C, Nieto JJ, Canovas M: Ectoines in cell stress protection: uses and biotechnological production. Biotechnol Adv 2010, 28:782-801. | ||
| In article | View Article PubMed | ||
| [9] | DAVID CA´NOVAS, 2, † NUNO BORGES CV, 1 JOAQUI´N J. NIETO,1, ANTONIO VENTOSA AHS: Role of Nγ-Acetyldiaminobutyrate as an Enzyme Stabilizer and an Intermediate in the Biosynthesis of Hydroxyectoine. 1999. | ||
| In article | |||
| [10] | Ning Y, Wu X, Zhang C, Xu Q, Chen N, Xie X: Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metab Eng 2016, 36:10-8. | ||
| In article | View Article PubMed | ||
| [11] | Zhao Q, Li S, Lv P, Sun S, Ma C, Xu P, Su H, Yang C: High ectoine production by an engineered Halomonas hydrothermalis Y2 in a reduced salinity medium. Microb Cell Fact 2019, 18:184. | ||
| In article | View Article PubMed | ||
| [12] | Dong Y, Zhang H, Wang X, Ma J, Lei P, Xu H, Li S: Enhancing ectoine production by recombinant Escherichia coli through step-wise fermentation optimization strategy based on kinetic analysis. Bioprocess Biosyst Eng 2021, 44:1557-66. | ||
| In article | View Article PubMed | ||
| [13] | He Y-Z, Gong J, Yu H-Y, Tao Y, Zhang S, Dong Z-Y: High production of ectoine from aspartate and glycerol by use of whole-cell biocatalysis in recombinant Escherichia coli. Microbial Cell Factories 2015, 14. | ||
| In article | View Article PubMed | ||
| [14] | Grammann K, Volke A, Kunte HJ: New type of osmoregulated solute transporter identified in halophilic members of the bacteria domain: TRAP transporter TeaABC mediates uptake of ectoine and hydroxyectoine in Halomonas elongata DSM 2581(T). J Bacteriol 2002, 184:3078-85. | ||
| In article | View Article PubMed | ||
| [15] | Jiang Y, Chen B, Duan C, Sun B, Yang J, Yang S: Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl Environ Microbiol 2015, 81:2506-14. | ||
| In article | View Article PubMed | ||
| [16] | HISAYO ONO KS, NONPANGA KHUNAJAKR,† TAO TAO,‡ MIHOKO YAMAMOTO, MASAYUKI HIRAMOTO, ATSUHIKO SHINMYO,§ MITSUO TAKANO, AND YOSHIKATSU MUROOKA: Characterization of Biosynthetic Enzymes for Ectoine as a Compatible Solute in a Moderately Halophilic Eubacterium, Halomonas elongata. 1999. | ||
| In article | |||
| [17] | Moritz1 LBKD: Boosting Escherichia coli’s heterologous production rate of ectoines by exploiting the non‑halophilic gene cluster from Acidiphilium cryptum, 2020. | ||
| In article | |||
| [18] | Parwata IP, Wahyuningrum D, Suhandono S, Hertadi R: Heterologous Ectoine Production in Escherichia coli: Optimization Using Response Surface Methodology. Int J Microbiol 2019, 2019:5475361. | ||
| In article | View Article PubMed | ||
| [19] | Oesterle S, Gerngross D, Schmitt S, Roberts TM, Panke S: Efficient engineering of chromosomal ribosome binding site libraries in mismatch repair proficient Escherichia coli. Sci Rep 2017, 7:12327. | ||
| In article | View Article PubMed | ||
| [20] | Czech L, Hoppner A, Kobus S, Seubert A, Riclea R, Dickschat JS, Heider J, Smits SHJ, Bremer E: Illuminating the catalytic core of ectoine synthase through structural and biochemical analysis. Sci Rep 2019, 9:364. | ||
| In article | View Article PubMed | ||
| [21] | Guo H, Yang Y, Xue F, Zhang H, Huang T, Liu W, Liu H, Zhang F, Yang M, Liu C, Lu H, Zhang Y, Ma L: Effect of flexible linker length on the activity of fusion protein 4-coumaroyl-CoA ligase::stilbene synthase. Mol Biosyst 2017, 13:598-606. | ||
| In article | View Article PubMed | ||
| [22] | Ying X, Wang C, Shao S, Wang Q, Zhou X, Bai Y, Chen L, Lu C, Zhao M, Wang Z: Efficient Oxidation of Methyl Glycolate to Methyl Glyoxylate Using a Fusion Enzyme of Glycolate Oxidase, Catalase and Hemoglobin. Catalysts 2020, 10. | ||
| In article | View Article | ||
| [23] | Hillier HT, Altermark B, Leiros I: The crystal structure of the tetrameric DABA-aminotransferase EctB, a rate-limiting enzyme in the ectoine biosynthesis pathway. FEBS J 2020, 287:4641-58. | ||
| In article | View Article PubMed | ||
Published with license by Science and Education Publishing, Copyright © 2021 Yuanyuan Yang, Songmao Wang, Kechen Yu, Shiyi Xu, Mengzhu Liu, Jianyong Zheng and Wei Yuan
![]() This work is licensed under a Creative Commons Attribution 4.0 International License. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/
This work is licensed under a Creative Commons Attribution 4.0 International License. To view a copy of this license, visit
http://creativecommons.org/licenses/by/4.0/
| [1] | Erwin A. GALINSKI’ H-PPaHGT: 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxy~c acid A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. 1985. | ||
| In article | View Article PubMed | ||
| [2] | Chen WC, Hsu CC, Lan JC, Chang YK, Wang LF, Wei YH: Production and characterization of ectoine using a moderately halophilic strain Halomonas salina BCRC17875. J Biosci Bioeng 2018, 125:578-84. | ||
| In article | View Article PubMed | ||
| [3] | Chen W, Zhang S, Jiang P, Yao J, He Y, Chen L, Gui X, Dong Z, Tang SY: Design of an ectoine-responsive AraC mutant and its application in metabolic engineering of ectoine biosynthesis. Metab Eng 2015, 30:149-55. | ||
| In article | View Article PubMed | ||
| [4] | Judith Becker1† RS, Michael Kohlstedt1, Björn J Harder1, Nicole S Borchert1, Nadine Stöveken2,3, Erhard Bremer2,3 and Christoph Wittmann1*: Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. 2013. | ||
| In article | |||
| [5] | Liu M, Liu H, Shi M, Jiang M, Li L, Zheng Y: Microbial production of ectoine and hydroxyectoine as high-value chemicals. Microb Cell Fact 2021, 20:76. | ||
| In article | View Article PubMed | ||
| [6] | Schwibbert K, Marin-Sanguino A, Bagyan I, Heidrich G, Lentzen G, Seitz H, Rampp M, Schuster SC, Klenk HP, Pfeiffer F, Oesterhelt D, Kunte HJ: A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581 T. Environ Microbiol 2011, 13:1973-94. | ||
| In article | View Article PubMed | ||
| [7] | Giesselmann G, Dietrich D, Jungmann L, Kohlstedt M, Jeon EJ, Yim SS, Sommer F, Zimmer D, Muhlhaus T, Schroda M, Jeong KJ, Becker J, Wittmann C: Metabolic Engineering of Corynebacterium glutamicum for High-Level Ectoine Production: Design, Combinatorial Assembly, and Implementation of a Transcriptionally Balanced Heterologous Ectoine Pathway. Biotechnol J 2019, 14:e1800417. | ||
| In article | View Article PubMed | ||
| [8] | Pastor JM, Salvador M, Argandona M, Bernal V, Reina-Bueno M, Csonka LN, Iborra JL, Vargas C, Nieto JJ, Canovas M: Ectoines in cell stress protection: uses and biotechnological production. Biotechnol Adv 2010, 28:782-801. | ||
| In article | View Article PubMed | ||
| [9] | DAVID CA´NOVAS, 2, † NUNO BORGES CV, 1 JOAQUI´N J. NIETO,1, ANTONIO VENTOSA AHS: Role of Nγ-Acetyldiaminobutyrate as an Enzyme Stabilizer and an Intermediate in the Biosynthesis of Hydroxyectoine. 1999. | ||
| In article | |||
| [10] | Ning Y, Wu X, Zhang C, Xu Q, Chen N, Xie X: Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metab Eng 2016, 36:10-8. | ||
| In article | View Article PubMed | ||
| [11] | Zhao Q, Li S, Lv P, Sun S, Ma C, Xu P, Su H, Yang C: High ectoine production by an engineered Halomonas hydrothermalis Y2 in a reduced salinity medium. Microb Cell Fact 2019, 18:184. | ||
| In article | View Article PubMed | ||
| [12] | Dong Y, Zhang H, Wang X, Ma J, Lei P, Xu H, Li S: Enhancing ectoine production by recombinant Escherichia coli through step-wise fermentation optimization strategy based on kinetic analysis. Bioprocess Biosyst Eng 2021, 44:1557-66. | ||
| In article | View Article PubMed | ||
| [13] | He Y-Z, Gong J, Yu H-Y, Tao Y, Zhang S, Dong Z-Y: High production of ectoine from aspartate and glycerol by use of whole-cell biocatalysis in recombinant Escherichia coli. Microbial Cell Factories 2015, 14. | ||
| In article | View Article PubMed | ||
| [14] | Grammann K, Volke A, Kunte HJ: New type of osmoregulated solute transporter identified in halophilic members of the bacteria domain: TRAP transporter TeaABC mediates uptake of ectoine and hydroxyectoine in Halomonas elongata DSM 2581(T). J Bacteriol 2002, 184:3078-85. | ||
| In article | View Article PubMed | ||
| [15] | Jiang Y, Chen B, Duan C, Sun B, Yang J, Yang S: Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl Environ Microbiol 2015, 81:2506-14. | ||
| In article | View Article PubMed | ||
| [16] | HISAYO ONO KS, NONPANGA KHUNAJAKR,† TAO TAO,‡ MIHOKO YAMAMOTO, MASAYUKI HIRAMOTO, ATSUHIKO SHINMYO,§ MITSUO TAKANO, AND YOSHIKATSU MUROOKA: Characterization of Biosynthetic Enzymes for Ectoine as a Compatible Solute in a Moderately Halophilic Eubacterium, Halomonas elongata. 1999. | ||
| In article | |||
| [17] | Moritz1 LBKD: Boosting Escherichia coli’s heterologous production rate of ectoines by exploiting the non‑halophilic gene cluster from Acidiphilium cryptum, 2020. | ||
| In article | |||
| [18] | Parwata IP, Wahyuningrum D, Suhandono S, Hertadi R: Heterologous Ectoine Production in Escherichia coli: Optimization Using Response Surface Methodology. Int J Microbiol 2019, 2019:5475361. | ||
| In article | View Article PubMed | ||
| [19] | Oesterle S, Gerngross D, Schmitt S, Roberts TM, Panke S: Efficient engineering of chromosomal ribosome binding site libraries in mismatch repair proficient Escherichia coli. Sci Rep 2017, 7:12327. | ||
| In article | View Article PubMed | ||
| [20] | Czech L, Hoppner A, Kobus S, Seubert A, Riclea R, Dickschat JS, Heider J, Smits SHJ, Bremer E: Illuminating the catalytic core of ectoine synthase through structural and biochemical analysis. Sci Rep 2019, 9:364. | ||
| In article | View Article PubMed | ||
| [21] | Guo H, Yang Y, Xue F, Zhang H, Huang T, Liu W, Liu H, Zhang F, Yang M, Liu C, Lu H, Zhang Y, Ma L: Effect of flexible linker length on the activity of fusion protein 4-coumaroyl-CoA ligase::stilbene synthase. Mol Biosyst 2017, 13:598-606. | ||
| In article | View Article PubMed | ||
| [22] | Ying X, Wang C, Shao S, Wang Q, Zhou X, Bai Y, Chen L, Lu C, Zhao M, Wang Z: Efficient Oxidation of Methyl Glycolate to Methyl Glyoxylate Using a Fusion Enzyme of Glycolate Oxidase, Catalase and Hemoglobin. Catalysts 2020, 10. | ||
| In article | View Article | ||
| [23] | Hillier HT, Altermark B, Leiros I: The crystal structure of the tetrameric DABA-aminotransferase EctB, a rate-limiting enzyme in the ectoine biosynthesis pathway. FEBS J 2020, 287:4641-58. | ||
| In article | View Article PubMed | ||



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}